뉴스브리핑센터
한의약융합데이터 콘텐츠 바로가기
동향분석
Home > 뉴스브리핑센터 > 동향분석


| NATURE에 실린 코로나19 (COVID-19) 아티클 소개 (~3/31) |
|
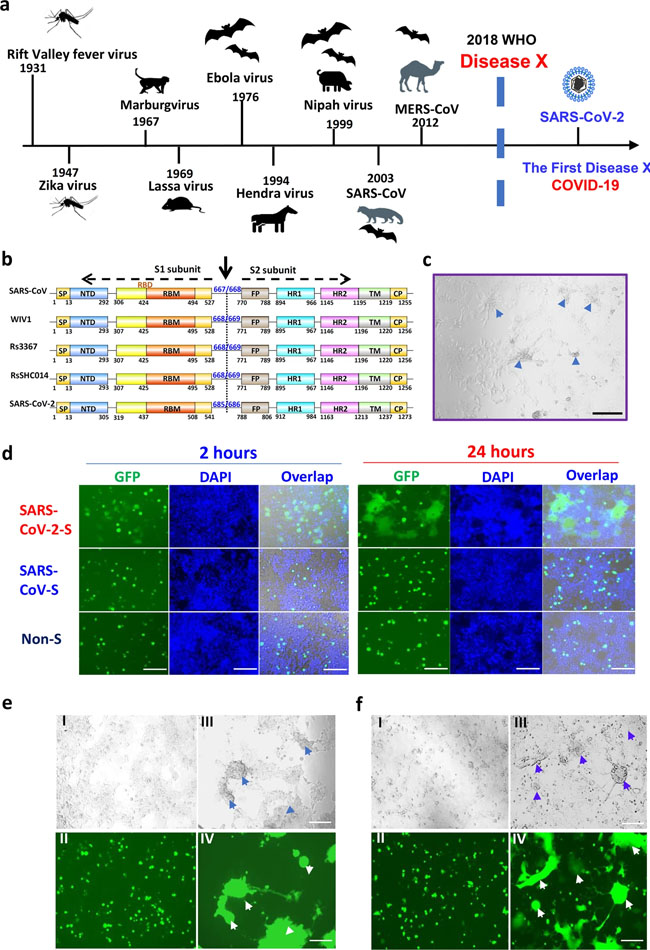
Comments and Opinion | 31 March 2020 코로나바이러스: 인류의 새로운 도전 COVID-19: a new challenge for human beings https://www.nature.com/articles/s41423-020-0407-x Correspondence | 30 March 2020 기저 질환으로 고혈압이 있는 코로나19 환자의 ACEI/ARB 항고혈압제 치료 Antihypertensive treatment with ACEI/ARB of patients with COVID-19 complicated by hypertension https://www.nature.com/articles/s41440-020-0433-1 Correspondence | 30 March 2020 코로나19에서 RAAS 억제제와 ACE2 간의 상호작용 Interaction between RAAS inhibitors and ACE2 in the context of COVID-19 https://www.nature.com/articles/s41569-020-0368-x News | 30 March 2020 우리는 조심해야 한다: 과학자들은 중국의 폐쇄가 풀리면서 제2의 코로나바이러스 사태를 우려 각국은 여행 제한을 해제하고 있는 후베이성의 재감염을 주시할 것이다. ‘We need to be alert’: Scientists fear second coronavirus wave as China’s lockdowns ease Other countries on lockdown will be watching for a resurgence of infections in Hubei province now that travel restrictions are lifting. https://www.nature.com/articles/d41586-020-00938-0 Research | 30 March 2020 막 융합을 중재할 수 있는 고용량 스파이크 단백질을 타겟으로 하는 매우 강력한 판 코로나바이러스 융합 억제제에 의한 SARS -CoV-2 (2019-nCoV) 감염 억제 Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion https://www.nature.com/articles/s41422-020-0305-x Fig. 1: Establishment of SARS-CoV-2 S protein-mediated cell–cell fusion system.
a The emerging timeline for highly pathogenic viruses and the proposed Disease X. b Schematic representation of SARS-CoV-2 S protein. Its S1 subunit contains NTD (14–305 aa), RBD (319–541 aa), and RBM (437–508 aa). Its S2 subunit contains FP (788–806 aa), HR1 (912–984 aa), HR2 (1163–1213 aa), TM (1214–1237 aa) and CP (1238–1273 aa). c The formation of syncytium in Huh-7 cells 24 h after SARS-CoV-2 infection, with scale bar of 200 µm. d Images of SARS-CoV and SARS-CoV-2 S-mediated cell–cell fusion on 293T/ACE2 cells at 2 h (left) and 24 h (right). e SARS-CoV (I–II) and SARS-CoV-2 (III–IV) S-mediated syncytium formation on 293T/ACE2 cells at 48 h. f SARS-CoV (I–II) and SARS-CoV-2 (III–IV) S-mediated syncytium formation on Huh-7 cells at 48 h. Scale bar equals 400 µm in d–f. Fig. 2: Overall structure of post-fusion 6-HB in SARS-CoV-2.
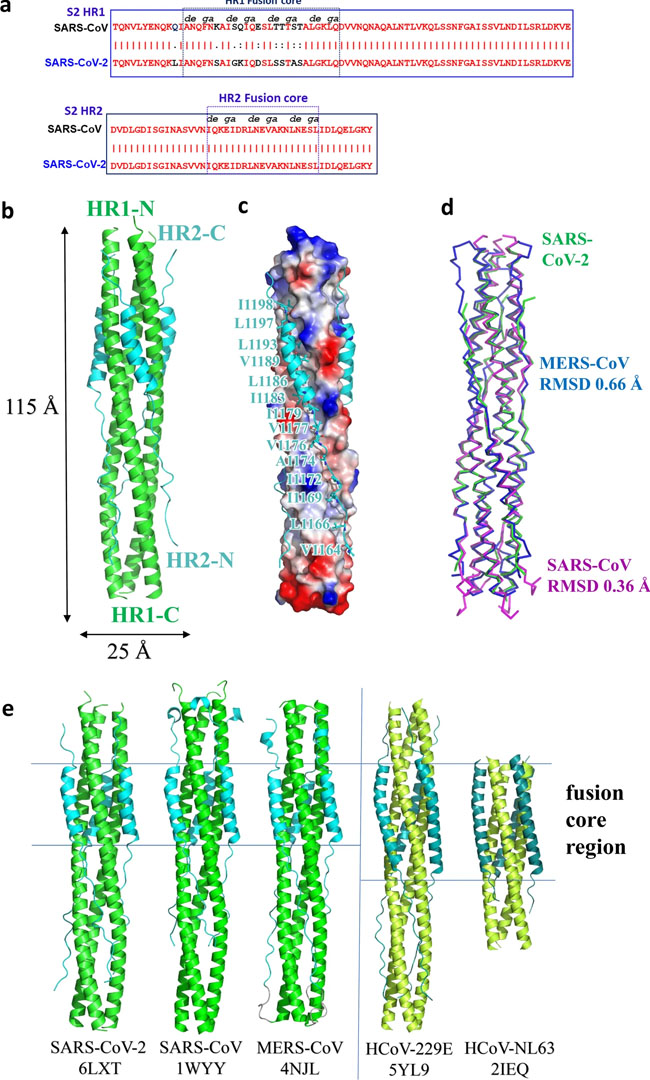
a Sequence alignment of HR1 and HR2 domains in SARS-CoV and SARS-CoV-2. b Structure of SARS-CoV-2 6-HB is shown in cartoon representation with HR1 colored in green and HR2 in cyan. The structural dimensions are indicated in angstroms. c HR1 trimer of SARS-CoV-2 6-HB is shown in electrostatic surface, and HR2 domain is shown in cartoon representation, the important binding residues of which are shown in sticks and labeled. d The superposition of 6-HB structure of SARS-CoV (PDB entry 1WYY), MERS-CoV (PDB entry 4NJL) and SARS-CoV-2 is shown in ribbon. The RMSD between structures is indicated. e The sequence comparison of 6-HB structure of different HCoVs is shown in cartoon representation with different colors for HR1 and HR2. The helical fusion core regions are indicated. Fig. 3: Interaction between HR1 and HR2 of SARS-CoV-2 and SARS-CoV.
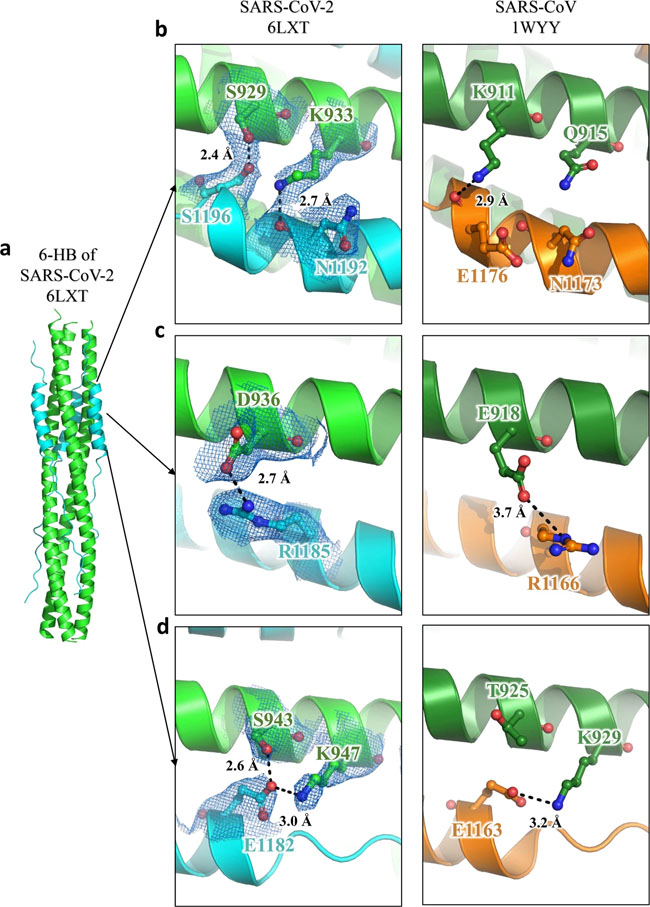
a–d The 6-HB structure of SARS-CoV-2 and SARS-CoV is shown in cartoon representation. The HR1 domain is shown in green for SARS-CoV-2 and forest for SARS-CoV, while the HR2 domain is shown in cyan for SARS-CoV-2 and orange for SARS-CoV. Important residues are shown in sticks and labeled. Fig. 4: EK1-Lipopeptides showed potent inhibitory activity against SARS-CoV-2 infection.
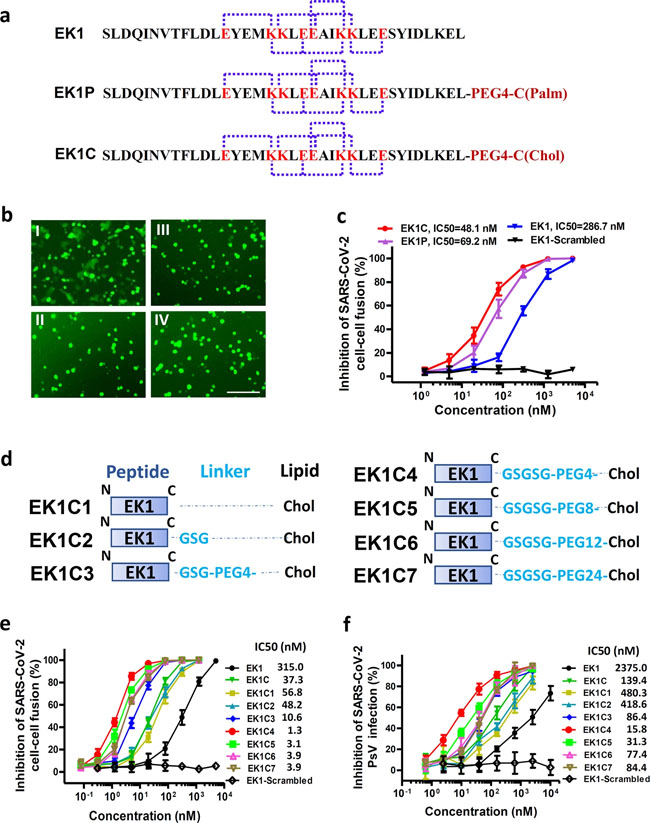
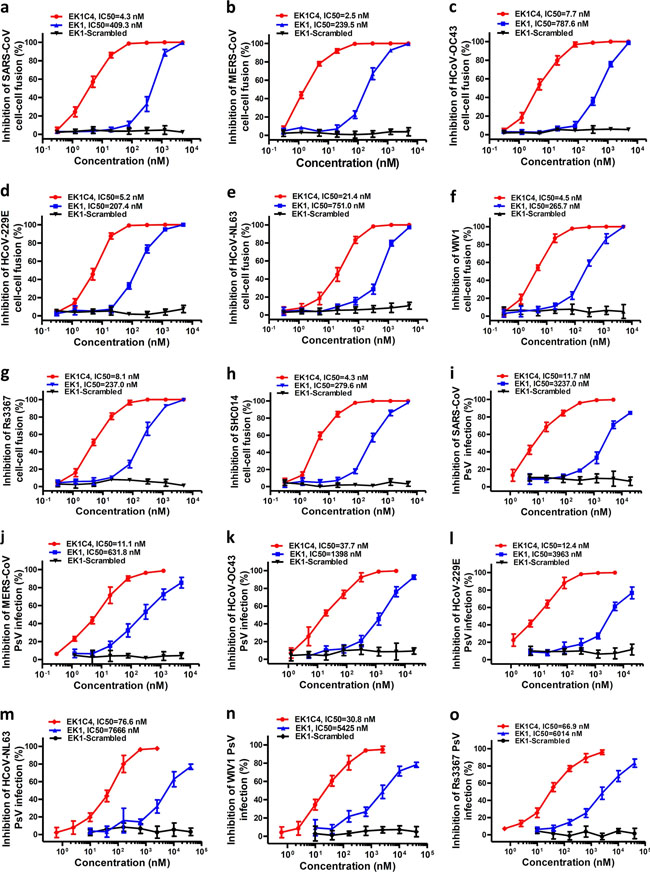
a Amino acid sequences of the designed peptides EK1, EK1P and EK1C. The dotted lines represent E–K salt-bridge with i to i + 3, or i + 4 arrangement. b SARS-CoV-2 S protein-mediated cell–cell fusion in the presence of EK1-scramble (I), EK1 (II), EK1C (III), and EK1P (IV) at 2.5 μM (scale bar: 400 µm). c Inhibitory activity of EK1-scramble, EK1, EK1C and EK1P against SARS-CoV-2 S-mediated cell–cell fusion. d Design diagram of EK1-lipopeptides with cholesterol modification, including EK1C1-EK1C7. e Inhibitory activity of EK1-lipopeptides on SARS-CoV-2 S-mediated cell–cell fusion. f Inhibitory activity of EK1-lipopeptides on SARS-CoV-2 PsV infection. Experiments were repeated twice, and the data are expressed as means ± SD (error bar). Fig. 5: EK1C4 broadly and potently inhibited cell–cell fusion and PsV infection mediated by S protein of divergent HCoVs.
a–h Inhibitory activity of EK1C4 in cell–cell fusion mediated by the S proteins of SARS-CoV (a), MERS-CoV (b), HCoV-OC43 (c), HCoV-229E (d), HCoV-NL63 (e), WIV1 (f), Rs3367 (g) and SHC014 (h). i–o Inhibitory activity of EK1C4 in PsV infection assays against SARS-CoV (i), MERS-CoV (j), HCoV-OC43 (k), HCoV-229E (l), NL63 (m), WIV1 (n) and Rs3367 (o). Experiments were repeated twice, and the data are expressed as means ± SD. Fig. 6: EK1C4 effectively inhibited live-CoVs infection in vitro and in vivo.
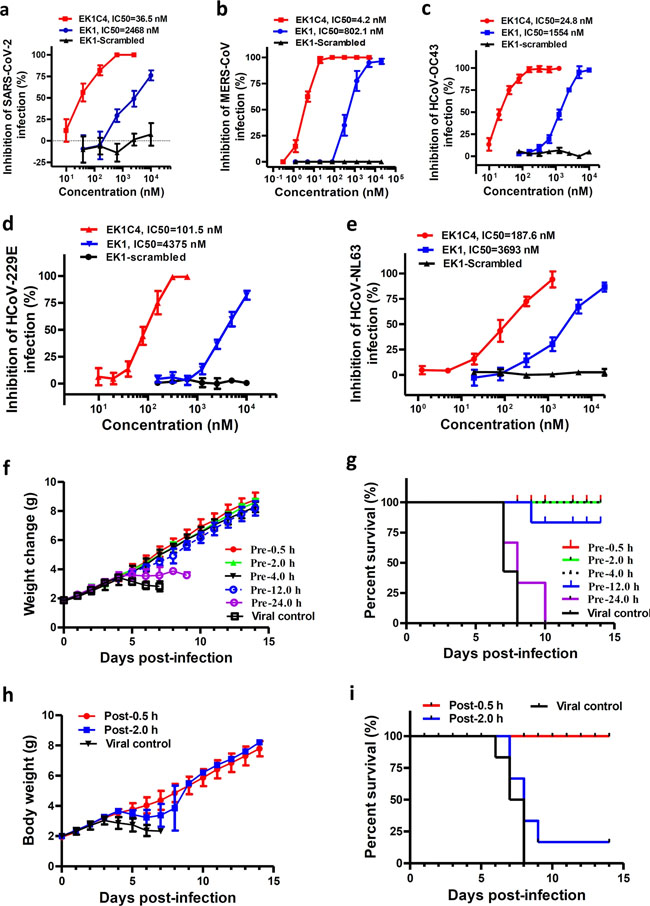
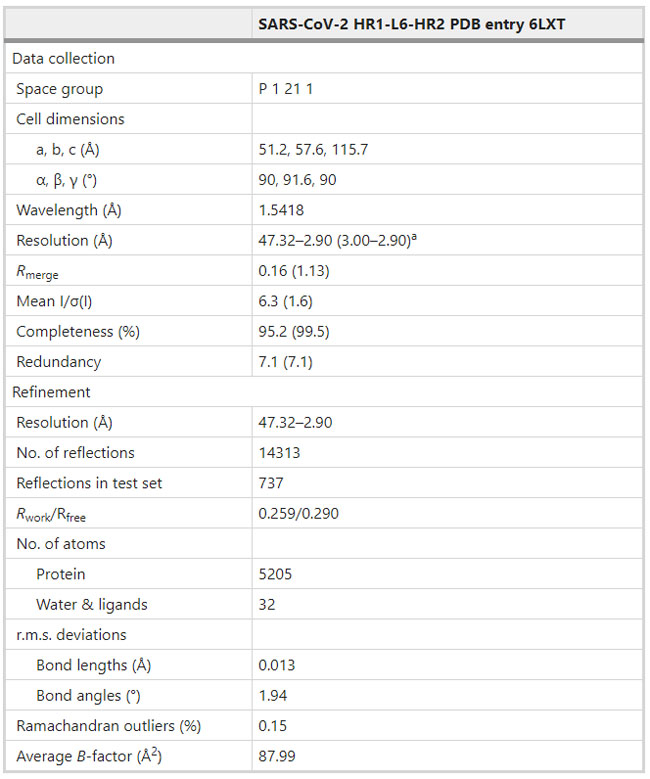
a–e Inhibitory activity of EK1C4 on live HCoV replication for SARS-CoV-2 (a), MERS-CoV (b), HCoV-OC43 (c), HCoV-229E (d), and HCoV-NL63 (e). f–g In vivo prophylactic efficacy of EK1C4 against HCoV-OC43 infection in mice. Body weight change (f) and survival curves (g) of mice challenged with HCoV-OC43. h–i In vivo therapeutic efficacy of EK1C4 against HCoV-OC43 infection in mice. Body weight change (h) and survival curves (i) of mice challenged with HCoV-OC43. Experiments were repeated twice, and the data are expressed as means ± SD. Table 1 Data collection and refinement statistics.
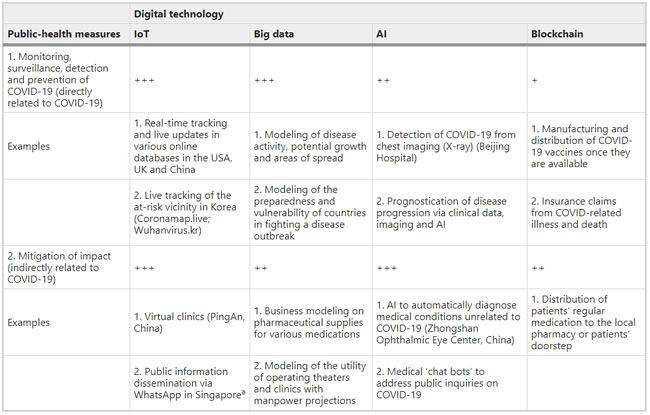
a: Highest resolution shell is shown in parenthesis. Comments and Opinion | 27 March 2020 디지털 기술과 코로나19 Digital technology and COVID-19 https://www.nature.com/articles/s41591-020-0824-5 Table 1 Digital technologies and their impact on public-health strategies
The likely impact of digital technologies on (1) disease monitoring, surveillance, detection and diagnosis, and (2) mitigation of impact: +, low (no clear example yet in either official government website); ++, moderate (one clear example); +++, high (two or more examples). Gray shading indicates potential applications that are not described in the literature thus far but should be considered by technology companies or research groups worldwide to help battle against COVID-19. Additional examples beyond those mentioned in the text are included in this table. Research | 27 March 2020 | OPEN 림프구 감소증은 COVID-19의 질병의 심각성을 예측한다: 기술적 예측 연구 Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study https://www.nature.com/articles/s41392-020-0148-4 Fig. 1
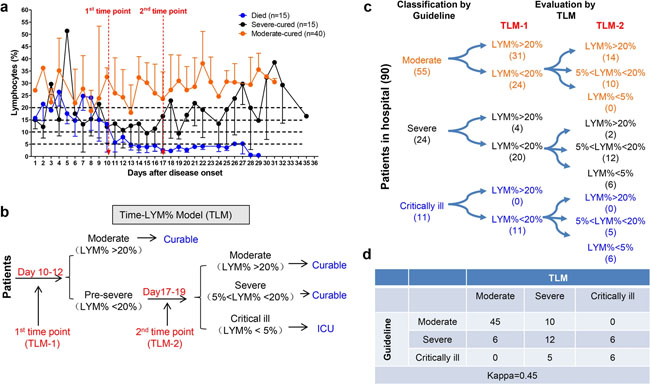
Establishment and validation of Time-LYM% model (TLM) in patients with COVID-19. a Dynamic changes of LYM% in the death cases (n = 15), severe-cured cases (n = 15), and moderate-cured cases (n = 40). Data are showen as means ± s.e.ms. Two cutoff time points of these three curves were set as 1st time point (day 10–12) and 2nd time point (day 17–19). b Description of TLM: 1st time point (TLM-1) and 2nd time point (TLM-2) are defined as day 10–12 and day 17–19 from symptom onset, respectively. The confirmed COVID-19 patients with LYM% > 20% at TLM-1 are classified as moderate type and the ones with LYM% < 20% at TLM-1 are suggested as pre-severe type, which need to be further distinguished at TLM-2. If LYM% > 20% at TLM-2, those pre-severe patients are reclassified as moderate. If 5% < LYM% < 20% at TLM-2, the pre-severe patients are indeed typed as severe. If LYM% < 5% at TLM-2, those patients are suggested as critically ill. The moderate and severe types are curable, while the critically ill types need intensive care has a poor prognosis. c Ninety COVID-19 patients were currently hospitalized in light of the classification criteria of the New Coronavirus Pneumonia Diagnosis Program (5th edition): 55 patients with moderate type, 24 patients with severe type and 11 patients with critically ill type. At TLM-1, LYM% in 24 out of 55 moderate cases was lower than 20%; At TLM-2, LYM% in all 24 patients was above 5%, indicating that these patients would be curable. Regarding other 24 patients with severe symptoms, LYM% at TLM-1 was lower than 20% in 20 out of 24 cases. LYM% at TLM-2 in 6 cases was <5%, indicating a poor prognosis. In 11 out of 11 critically ill patients, LYM% at TLM-1 was lower than 20%. LYM% at TLM-2 in six cases was lower than 5%, suggesting a poor prognosis. d The consistency between Guideline and TLM-based disease classification in c was tested using kappa statistic. Kappa = 0.48; P < 0.005 Research | 27 March 2020 SARS-CoV-2의 스파이크 당단백질 특성 및 SARS-CoV와의 면역 상호반응 Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV https://www.nature.com/articles/s41467-020-15562-9 Fig. 1: Incorporation of SARS-CoV-2 S protein into pseudovirions.
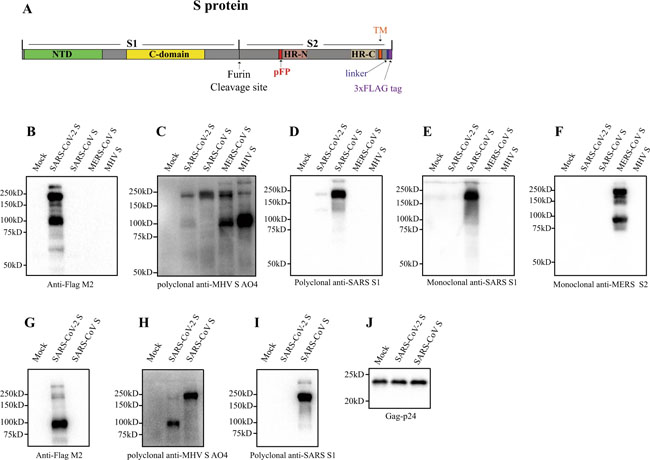
a Diagram of full-length SARS-CoV-2 S protein with a 3xFLAG tag. S1, receptor-binding subunit; S2, membrane fusion subunit; TM, transmembrane domain; NTD, N-terminal domain; pFP, potential fusion peptide; HR-N, heptad repeat-N; HR-C, heptad repeat-C; b–f Detection of CoVs S protein in cells lysate by western blot. Mock, 293T cells transfected with empty vector. b Mouse monoclonal anti-FLAG M2 antibody; c Polyclonal goat anti-MHV-A59 S protein antibody AO4. d Polyclonal rabbit anti-SARS S1 antibodies T62. e Mouse monoclonal anti-SARS S1 antibody. f Mouse monoclonal anti-MERS-CoV S2 antibody. g–j Detection of CoVs S protein in pseudovirions by western blot.Gag-p24 served as a loading control. g Anti-FLAG M2. h Polyclonal goat anti-MHV-A59 S protein antibody AO4. i Polyclonal rabbit anti-SARS S1 antibodies T62. j Polyclonal anti-Gag-p24 antibodies. uncleaved S protein, about 180 kDa; cleaved S protein, about 90 kDa. Experiments were done twice and one is shown. Source data are provided as a Source Data file. Fig. 2: Entry and receptor of SARS-CoV-2 S pseudovirons.
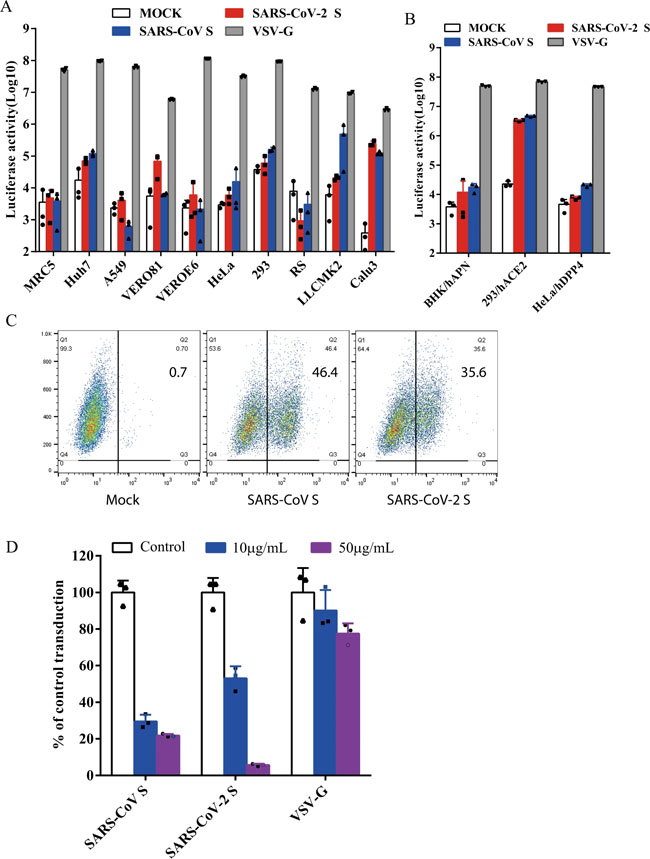
a, b Entry of SARS-CoV-2 S pseudovirions on indicated cell lines. Cells from human and animal origin were inoculated with SARS-CoV-2 S (red), SARS-CoV S (blue), or VSV-G (gray) pseudovirions. At 48 h post inoculation, transduction efficiency was measured according to luciferase activities. RS, Rhinolophus sinicus bat embryonic fibroblast; BHK/hAPN, BHK cells stably expressing hAPN, the hCoV-229E receptor; 293/hACE2, 293 cells stably expressing hACE2, the SARS-CoV receptor; HeLa/hDPP4, HeLa cells stably expressing hDPP4, the MERS-CoV receptor. Experiments were done in triplicates and repeated at least three times. One representative is shown with error bars indicating SEM. c Binding of SARS-CoV S and SARS-CoV-2 S proteins to soluble hACE2. HEK293T cells transiently expressing SARS-CoV and SARS-CoV-2 S proteins were incubated with the soluble hACE2 on ice, followed by polyclonal goat anti-hACE2 antibody. Cells were analyzed by flow cytometry. The experiments were repeated at least three times. d Inhibition of SARS-CoV-2 S pseudovirion entry by soluble hACE2. SARS-CoV S, SARS-CoV-2 S, or VSV-G pseudovirions were pre-incubated with soluble hACE2, then mixture were added to 293/hACE2 cells. Cells were lysed 40 h later and pseudoviral transduction was measured. Experiments were done twice and one representative is shown. Error bars indicate SEM of technical triplicates. Source data are provided as a Source Data file. Fig. 3: Endocytosis of SARS-CoV-2 S pseudovirions on 293/hACE2 cells.
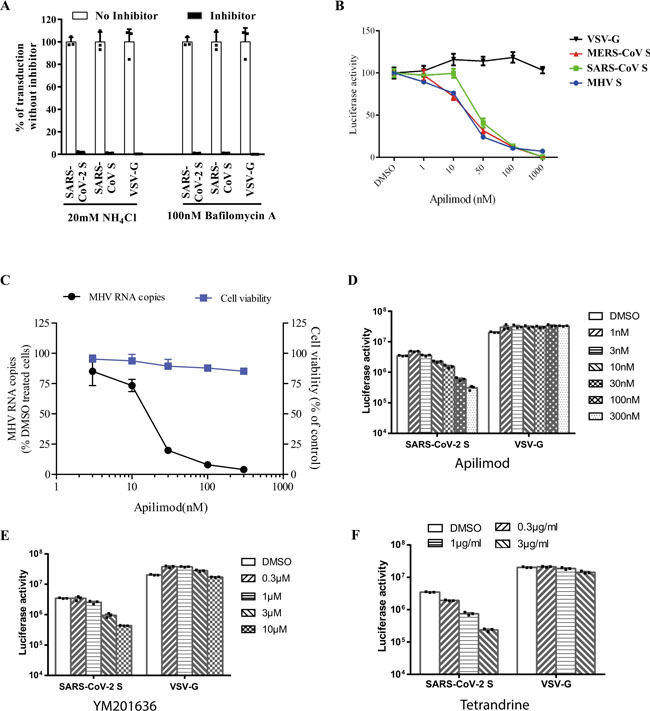
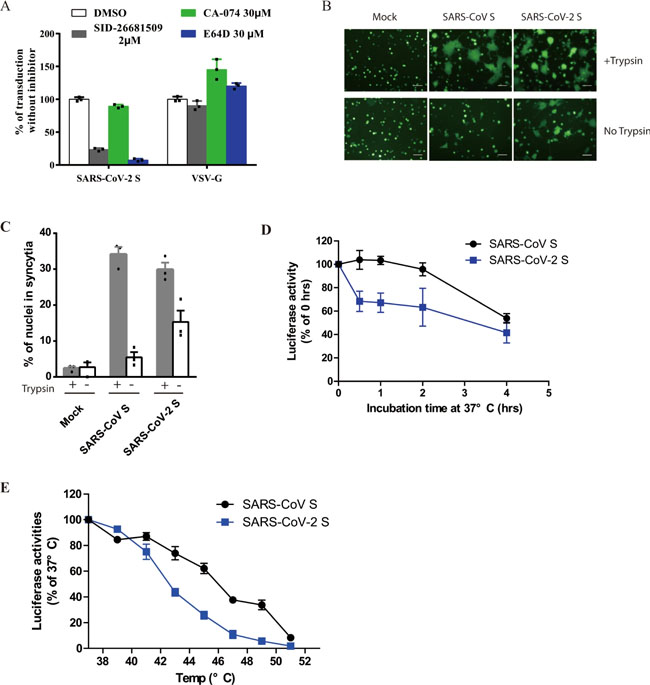
a Inhibition of entry of SARS-CoV-2 S pseudovirion on 293/hACE2 by lysosomotropic agents (20 mM NH4Cl and 100 nM bafilomycin A). b Inhibition of entry of SARS-CoV, MERS-CoV, and MHV S pseudovirions by a PIKfyve inhibitor apilimod. HeLa/mCEACAM, 293/hACE2, HeLa/hDPP4 cells were pretreated with different concentrations of apilimod and transduced with MHV S, SARS-CoV S, MERS-CoV S pseudovirions, respectively. The luciferase activity was measured 40 h post transduction. VSV-G pseudovirions were used as a control. Experiments were done in triplicates and repeated at least three times. One representative is shown with error bars indicating SEM. c Inhibition of MHV A59 infection by apilimod. The 17Cl.1 cells were pretreated with 3, 10, 30, 100, 300 nM apilimod for 30 min and infected by MHV A59 at MOI = 0.01. Viral infection and cell viability were determined by using qPCR and MTT assay, respectively. Experiments were done in triplicates and repeated at least three times. One representative is shown with error bars indicating SEM. d, e Inhibition of entry of SARS-CoV-2 S protein pseudovirions by apilimod, YM201636, and tetrandrine. HEK 293/hACE2 cells were pretreated with either apilimod (d), YM201636 (e), or tetrandrine (f), then inoculated with SARS-CoV-2 S pseudovirons in the presence of drug. The luciferase activity were measured 40 h post transduction. YM201636, PIKfyve inhibitor; tetrandrine, TPC2 inhibitor. The experiments were done in triplicates and repeated at least three times. One representative is shown with error bars indicating SEM of technical triplicates. Source data are provided as a Source Data file. Fig. 4: Activation of SARS-CoV-2 S protein by cathepsin and trypsin.
a Effects of cathepsin inhibitors on entry of SARS-CoV-2 S pseudovirions on 293/hACE2 cells. HEK 293/hACE2 cells were pretreated with broad-spectrum cathepsin inhibitor E64D, cathepsin L-specific inhibitor (SID 26681509), or cathepsin B-specific inhibitor (CA-074) and then transduced with SARS-CoV-2 S and VSV-G pseudovirions. Pseudoviral transduction was measured at 40 h post inoculation. Experiments were done in triplicates and repeated at least three times. One representative is shown. Error bars indicate SEM of technical triplicates. b Cell–cell fusion mediated by SARS-CoV-2 S protein. HEK 293T cells were transiently expressing eGFP and either SARS-CoV-2 or SARS-CoV S protein were detached with either trypsin or EDTA, and co-cultured with 293/hACE2 or 293 cells for 4 h at 37 °C. The scale bar indicates 250 µm. c Quantitative analysis of syncytia in panel b. d, e Thermostability analysis of SARS-CoV-2 S protein. d SARS-CoV and SARS-CoV-2 S pseudovirons were incubated at 37 °C for the specified times (0 to 4 h) in the absence of serum, and then assayed on 293/hACE2 cells. The results from infection at 0 h were set as 100%, and the experiments were repeated four times, and means with standard deviations are shown. e SARS-CoV and SARS-CoV-2 S pseudovirions without serum were incubated at the indicated temperature (37 to 51 °C) for 2 h and then assayed on 293/hACE2 cells. The results are reported as the percentage of transduction at 37 °C. The experiments were repeated four times, and means with standard deviations are shown. Source data are provided as a Source Data file. Fig. 5: Characterization of polyclonal rabbit anti-SARS S1 antibodies T62.
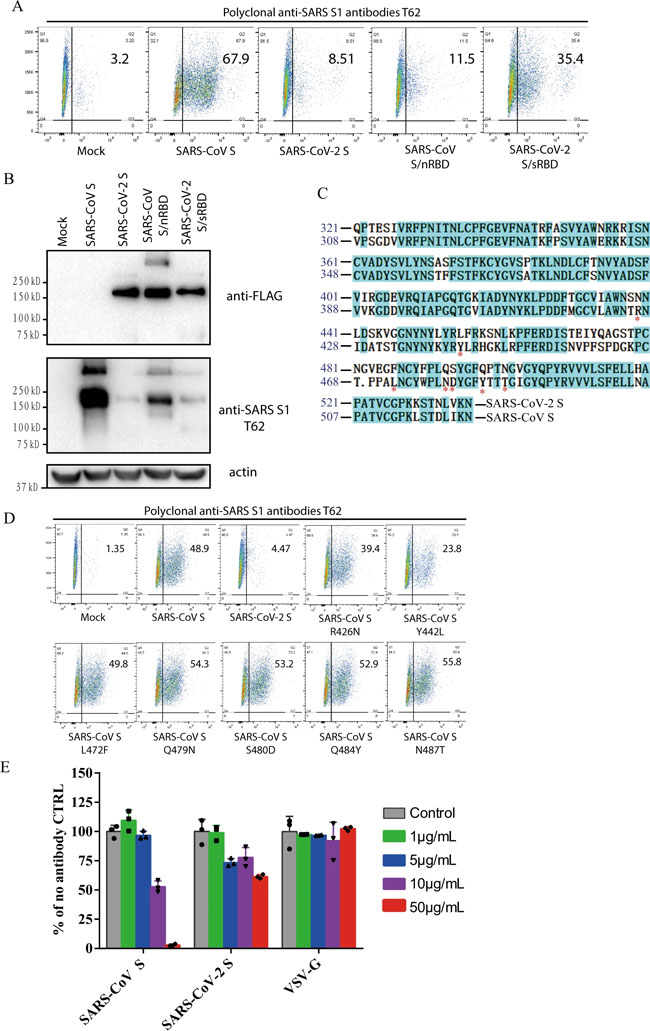
a Binding of polyclonal rabbit anti-SARS S1 antibodies T62 to SARS-CoV-2, SARS-CoV S, and chimeric S proteins. HEK293T cells transiently expressing either SARS-CoV-2 S, SARS-CoV S, SARS-CoV S/nRBD, or SARS-CoV-2 S/sRBD proteins were incubated with polyclonal rabbit anti-SARS-CoV S1 antibody T62 for 1 h on ice, followed by a FITC-conjugated secondary antibody, then cells were analyzed by flow cytometry. Experiments were done three times and one representative is shown. b Expression of SARS-CoV-2 S, SARS-CoV S, or chimeric S proteins on 293T cells. Cells from panel A were lyzed and blotted with anti-FLAG M2 antibody and polyclonal anti-SARS S1 antibody T62. c Amino acid sequence alignment of SARS-CoV and SARS-CoV-2 S RBDs. Stars (*) indicate the seven critical residues different between SARS-CoV-2 and SARS-CoV RBDs. d Binding of polyclonal rabbit anti-SARS S1 antibodies T62 to mutant SARS-CoV S proteins. e Neutralization of SARS-CoV-2 S and SARS-CoV S pseudovirions by polyclonal rabbit anti-SARS S1 antibody T62. Pseudovirons were pre-incubated with serially diluted polyclonal rabbit anti-SARS S1 antibodies T62 on ice, then virus-antibody mixture was added on 293/hACE2 cells. Pseudoviral transduction was measured 40 h later. Experiments were done in triplicates and repeated twice, and one representative is shown. Error bars indicate SEM of technical triplicates. Source data are provided as a Source Data file. Fig. 6: Limited cross-neutralization of SARS and COVID-19 sera.
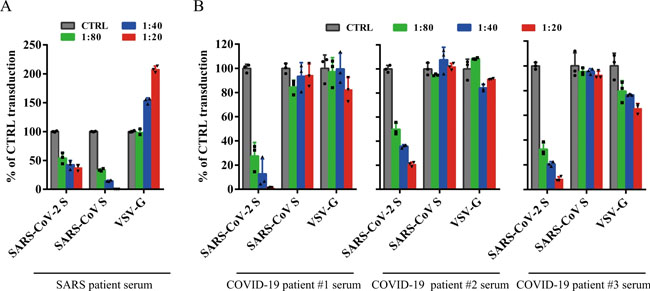
All sera were incubated on 56 °C for 30 min to eliminate complement. SARS-CoV S and SARS-CoV-2 S pseudovirons were pre-incubated with serially diluted SARS patient serum (a) or COVID-19 patient sera (b) for 1 h on ice and then added on 293/hACE2 cells. Pseudoviral transduction was measured 40 h later. Experiments were done in triplicates and repeated twice, and one representative is shown. Error bars indicate SEM of technical triplicates. Source data are provided as a Source Data file. Table 1 Neutralization activities of antisera from SARS-CoV and COVID-19 patients.
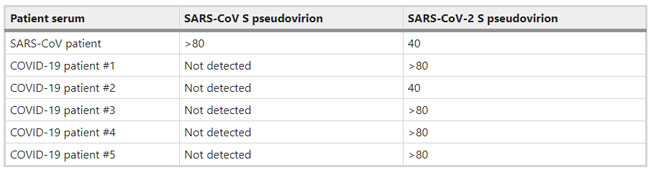
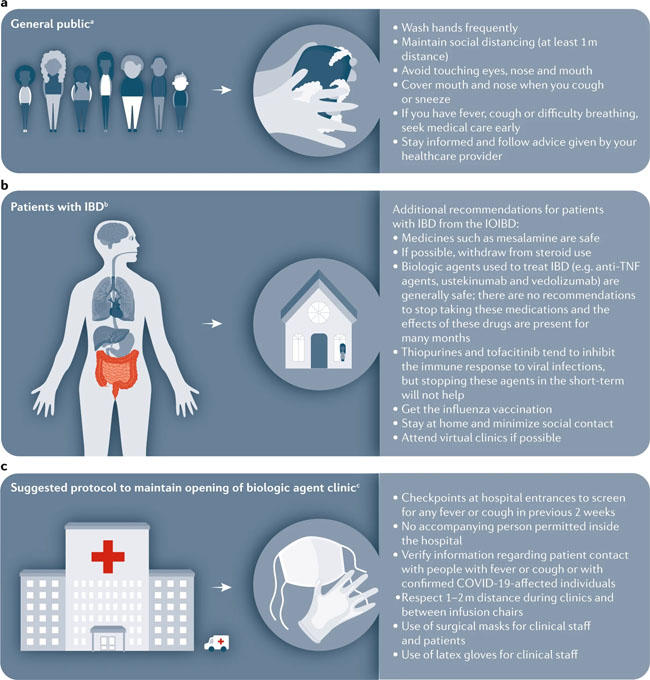
SARS-CoV and SARS-CoV-2 S pseudovirions were pre-incubated with serially diluted patient sera for 1 h on ice, then virus-antibody mixture was added on 293/hACE2 cells. Cells were lysed 40 h later and pseudovirus transduction was measured. The highest dilution of the serum sample that decreased transduction by 50% or more was considered to be positive. Source data are provided as a Source Data file. News | 27 March 2020 코로나바이러스 논문: 완치자의 혈장을 투여한 후 코로나19로 악화된 환자가 호전되다. Coronavirus papers: Debilitated patients rally after dose of survivors’ blood https://www.nature.com/articles/d41586-020-00502-w Comments and Opinion | 26 March 2020 코로나19 시기의 정신 건강 및 웰빙 관리를 위한 7가지 팁 자가격리와 갑작스러운 e-러닝 적응에 당황스러운 사람들을 위한 임상심리학자의 조언 Seven tips to manage your mental health and well-being during the COVID-19 outbreak Feeling overwhelmed by a lockdown and the need to suddenly adopt e-learning? Keep connected and compassionate, says clinical psychologist Desiree Dickerson. 1) Manage your expectations 2) Proactively manage your stress threshold 3) Know your red flags 4) Routine is your friend 5) Be compassionate with yourself and with others 6) Maintain connections 7) Manage uncertainty by staying in the present https://www.nature.com/articles/d41586-020-00933-5 News | 26 March 2020 다이아몬드 프린세스 크루즈선에서의 코로나19 감염 사례 폐쇄된 장소는 바이러스의 확산을 방지하지만 신종 코로나바이러스가 작용 기전 연구에 이상적 What the cruise-ship outbreaks reveal about COVID-19 Close confines help the virus to spread, but closed environments are also an ideal place to study how the new coronavirus behaves. https://www.nature.com/articles/d41586-020-00885-w News | 26 March 2020 과학자들은 백신을 테스트하기 위해 건강한 사람들을 코로나바이러스에 감염시켜야 하는가? 인간의 도전' 연구를 수행하자는 극단적인 제안은 백신 연구 속도에 박차를 가할 수 있다. Should scientists infect healthy people with the coronavirus to test vaccines? Radical proposal to conduct ‘human challenge’ studies could dramatically speed up vaccine research. https://www.nature.com/articles/d41586-020-00927-3 Comments and Opinion | 25 March 2020 코로나19 발생 시 염증성 장 질환 (inflammatory bowel disease, IBD) 관리: 임상 우선권 재설정 Management of IBD during the COVID-19 outbreak: resetting clinical priorities https://www.nature.com/articles/s41575-020-0294-8 Fig. 1: Approaches to minimize spread of infectious disease for patients with IBD during COVID-19.
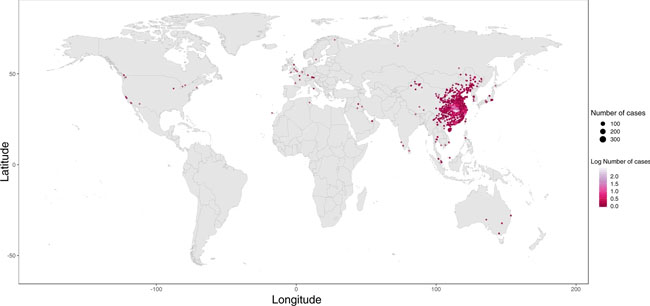
a | The WHO recommends basic protective measures against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) for the general public, including washing hands frequently and maintaining social distancing. b | The International Organization for the study of Inflammatory Bowel Disease (IOIBD) have made several recommendations for patients with inflammatory bowel disease (IBD) related to coronavirus disease (COVID-19), including the continuation of immunosuppressants. c | Strategies to enable maintenance of our biologic agent clinic during the COVID-19 outbreak in Italy include checkpoints at hospital entrances for symptom screening and use of surgical masks for clinical staff and patients. aBasic protective measures for the public according to the WHO. bRecommendations by IOIBD correct as of 16 Mar 2020. cExperience at the IBD Center, Humanitas University, Milan, Italy. News | 25 March 2020 코로나바이러스로 인해 여러 가지 다른 질병에 대한 약물 임상 시험 중단 의료기관 부족과 노출 위험에 대한 두려움이 임상연구에 제동을 걸면서 연구는 중단되었다. Coronavirus shuts down trials of drugs for multiple other diseases Studies grind to a halt as fears of health-care shortages and risk of exposure put the brakes on clinical research. https://www.nature.com/articles/d41586-020-00889-6 Research Highlights | 25 March 2020 코로나19: 대변-구강 경로 전염? COVID-19: faecal–oral transmission? https://www.nature.com/articles/s41575-020-0295-7 Correspondence | 24 March 2020 코로나19 위기 시 실험실 동물 관리 Care for laboratory animals during COVID-19 crisis https://www.nature.com/articles/d41586-020-00869-w News | 24 March 2020 코로나바이러스 완치자의 혈액이 생명을 구한다. How blood from coronavirus survivors might save lives https://www.nature.com/articles/d41586-020-00895-8 Research | 24 March 2020 코로나19 발병 역학 데이터, 실시간 확진자 정보 Epidemiological data from the COVID-19 outbreak, real-time case information https://www.nature.com/articles/s41597-020-0448-0 Fig. 1
Global distribution of reported confirmed cases from December 1, 2019 to February 5, 2020.
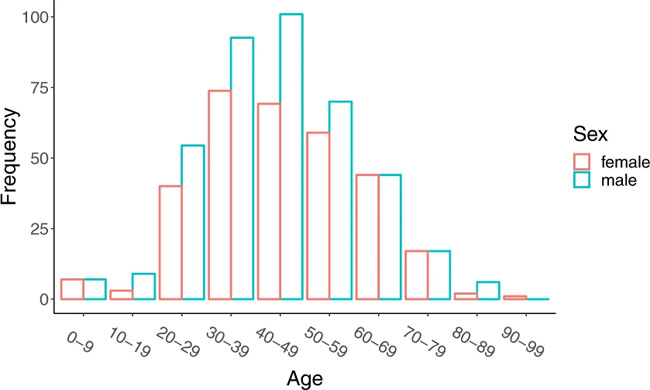
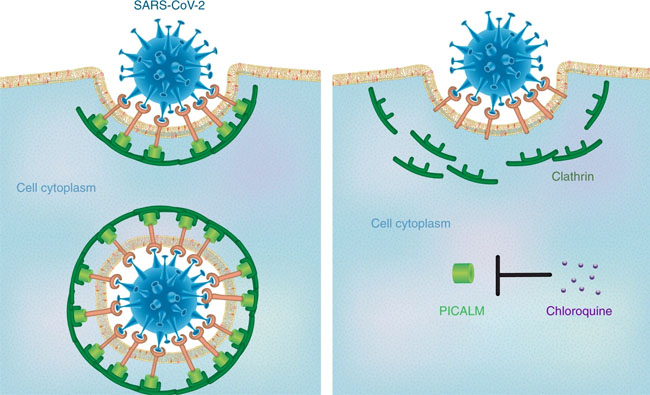
Age and sex distribution of confirmed cases globally (excluding Hubei). Comments and Opinion | 23 March 2020 나노의학의 관점에서 본 코로나19에 대한 클로로퀸의 효능 Insights from nanomedicine into chloroquine efficacy against COVID-19 https://www.nature.com/articles/s41565-020-0674-9 Fig. 1: Potential mechanism by which chloroquine exerts therapeutic effects against COVID-19.
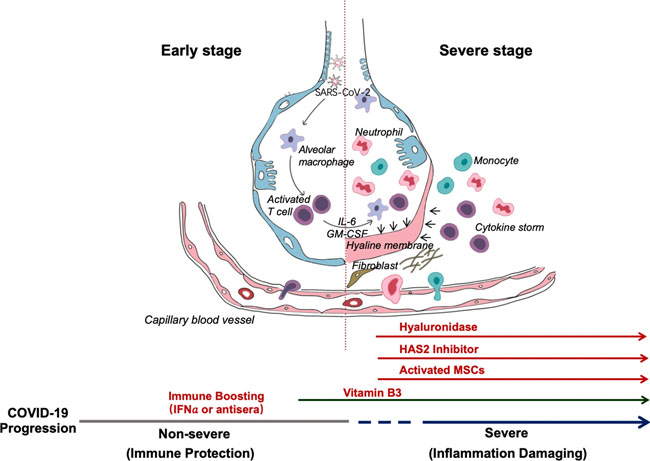
The proposed mechanism involves chloroquine-induced suppression of PICALM, which prevents endocytosis-mediated uptake of SARS-CoV-2. Editorial | 23 March 2020 코로나19 감염: 면역 반응에 대한 통찰력 COVID-19 infection: the perspectives on immune responses https://www.nature.com/articles/s41418-020-0530-3 Fig. 1: Schematic representation of the progression of COVID-19 infection and potential adjuvant interventions.
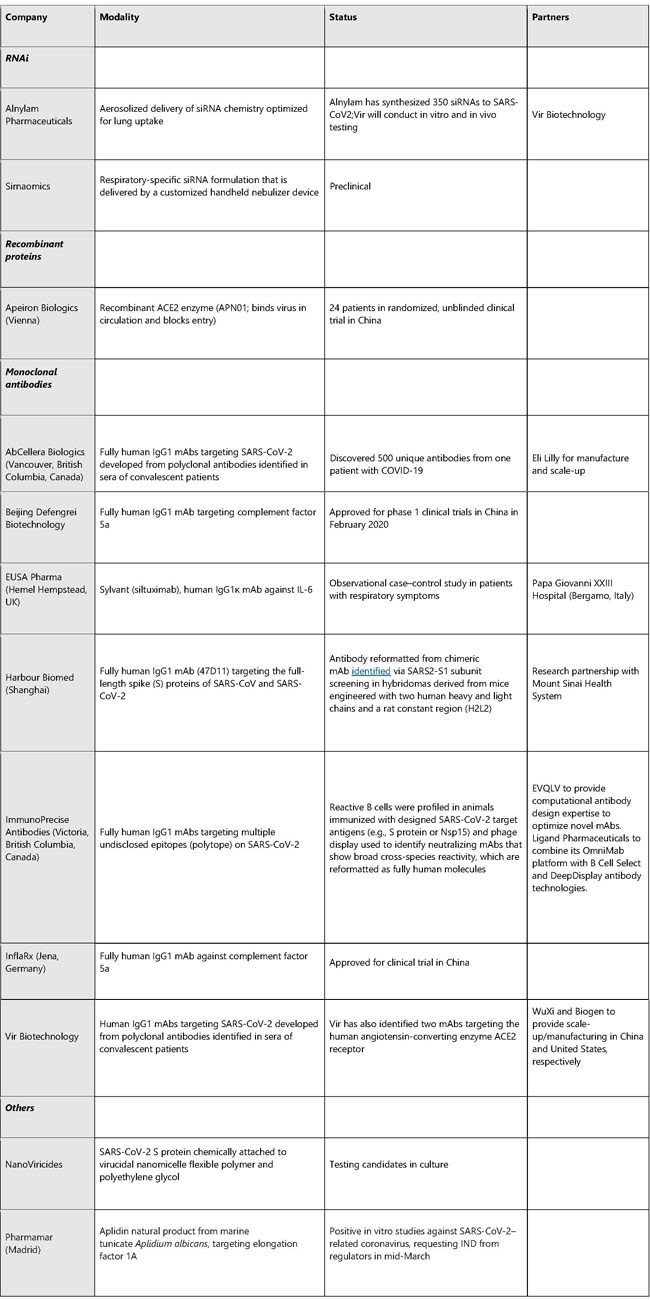
After an incubation period, the invading COVID-19 virus causes non-severe symptoms and elicits protective immune responses. The successful elimination of the infection relies on the health status and the HLA haplotype of the infected individual. In this period, strategies to boost immune response can be applied. If the general health status and the HLA haplotype of the infected individual do not eliminate the virus, the patient then enters the severe stage, when strong damaging inflammatory response occurs, especially in the lungs. At this stage, inhibition of hyaluronan synthase and elimination of hyaluronan can be prescribed. Cytokine activated mesenchymal stem cells can be used to block inflammation and promote tissue reparation. Vitamin B3 can be given to patients starting to have lung CT image abnormalities. News | 23 March 2020 코로나바이러스 팬데믹을 억제를 위한 빠르고 휴대가능한 진단 키트의 온라인 제공 진단 키트의 택배 배송과 디지털 장비들이 코로나19 확산을 막기 위해 갖춰졌다. Fast, portable tests come online to curb coronavirus pandemic Testing kits delivered by courier and digital tools combine to battle the COVID-19 outbreak. https://www.nature.com/articles/d41587-020-00010-2 Correspondence | 23 March 2020 COVID-19가 전 세계 조혈 줄기세포 기증자 제공에 미치는 영향: 부수적인 피해 The impact of COVID-19 on the provision of donor hematopoietic stem cell products worldwide: collateral damage https://www.nature.com/articles/s41409-020-0873-x News | 23 March 2020 코로나 검사: 연구자들은 팬데믹을 극복하기 위해 새로운 진단법을 찾는다. 네이처는 바이러스 진단 검사 진행 방법, 전 세계적으로 검사법이 다양한 이유 및 코로나19를 극복하기 위해 개발 중인 CRISPR 기반 검사에 대해 조사한다. Coronavirus tests: researchers chase new diagnostics to fight the pandemic Nature examines how viral diagnostic tests work, why testing has varied around the world and the CRISPR-based tests under development to fight COVID-19. https://www.nature.com/articles/d41586-020-00827-6 News | 20 March 2020 팬데믹 파이프라인 The pandemic pipeline https://www.nature.com/articles/d41587-020-00005-z TABLE 1. SELECTED EXPERIMENTAL THERAPIES UNDER DEVELOPMENT FOR COVID-19
Does not include polyclonal or IgG products extracted from convalescent patient serum. Sources: BioWorld, company sites, Thomson Cortellis, PubMed TABLE 2. SELECTED VACCINES UNDER DEVELOPMENT FOR COVID-19
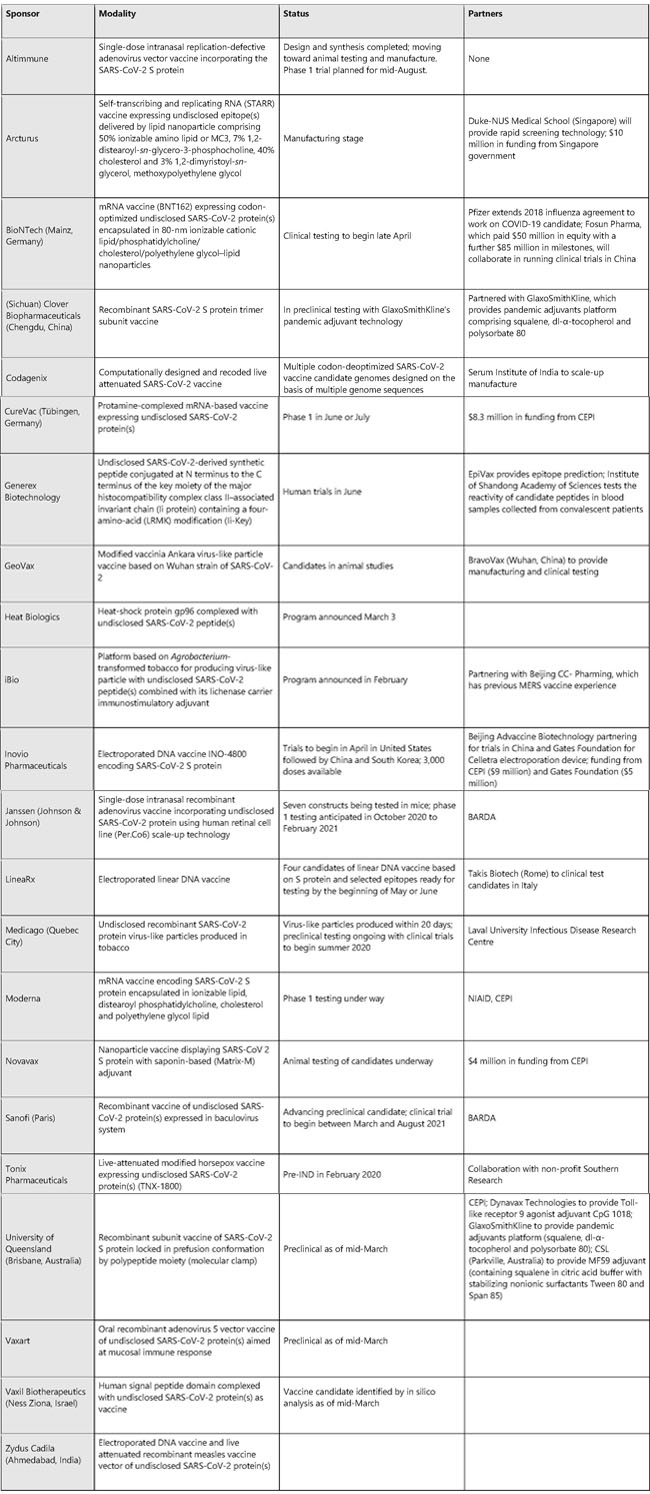
Sources: BioWorld, company sites, Thomsen Cortellis, PubMed News | 20 March 2020 진단, 진단, 진단! 코로나바이러스 발생을 막는 데 진단과 접촉 추적이 핵심이라면, 왜 전 세계에서 행해지고 있지 않는 것일까? Coronapod: “Test, test, test!” If testing and contact tracing are key to controlling the coronavirus outbreak, why aren’t they being done around the world? https://www.nature.com/articles/d41586-020-00864-1 News | 20 March 2020 코로나바이러스 확진자의 은폐로 새로운 감염이 퍼질 수 있다. 과학자들은 이 병원균에 감염된 경미한 증상이 있거나 증상이 없는 사람들의 비율을 추정하기 위해 박차를 가하고 있다. Covert coronavirus infections could be seeding new outbreaks Scientists are rushing to estimate the proportion of people with mild or no symptoms who could be spreading the pathogen. https://www.nature.com/articles/d41586-020-00822-x News | 20 March 2020 코로나바이러스에 당면한 과학자들의 의문: 왜 우리는 공지 받지 못했나? 미국 당국은 확산 대응의 초석인 진단과 접촉에 대한 정보 제공에 실패했다. Scientists exposed to coronavirus wonder: why weren’t we notified? US authorities are failing to test people and notify their contacts, a cornerstone of outbreak response. https://www.nature.com/articles/d41586-020-00823-w Research | 19 March 2020 중국 우한시 감염 양상으로 추정한 COVID-19의 임상 심각도 Estimating clinical severity of COVID-19 from the transmission dynamics in Wuhan, China https://www.nature.com/articles/s41591-020-0822-7 Fig. 1: Data used in the inference
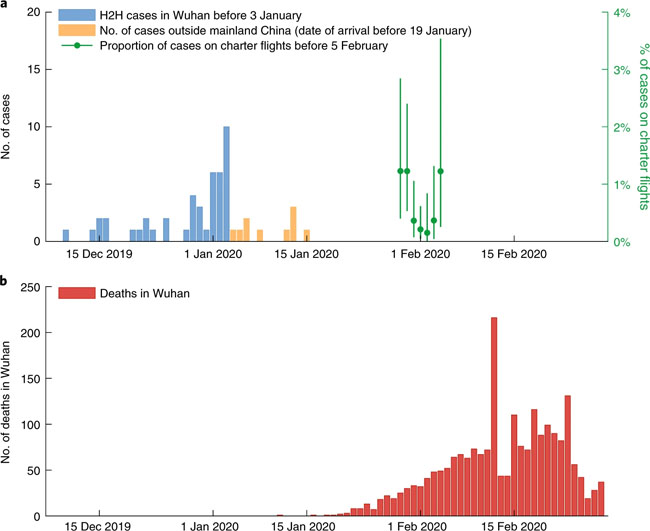
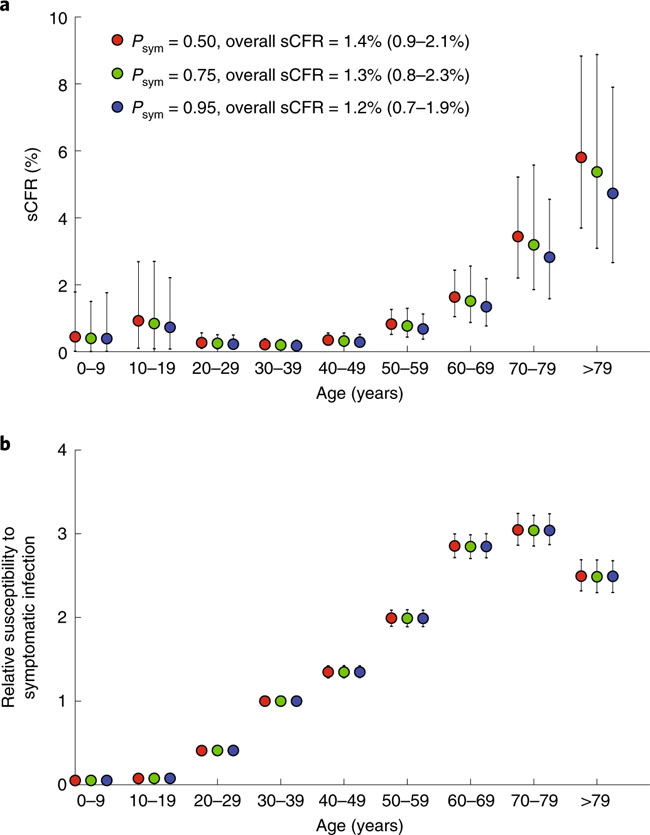
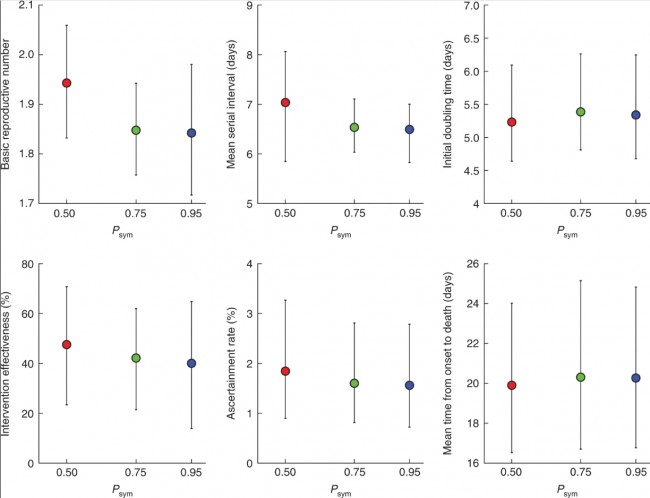
a, The daily number of confirmed cases in Wuhan (with no epidemiologic links to Huanan Seafood Wholesale Market, i.e., cases due to human-to-human (H2H) transmission) between 1 December 2019 and 3 January 2020 (blue), the daily number of cases exported from Wuhan to cities outside mainland China via air travel between 25 December 2019 and 19 January 2020 (orange) and the proportion of expatriates on charter flights between 29 January and 4 February 2020 who were laboratory-confirmed to be infected (green). The numbers of passengers and confirmed cases who returned to their countries from Wuhan on chartered flights are provided in Supplementary Table 3. Bars indicate the 95% confidence intervals (CIs) of the proportion. b, The daily number of deaths in Wuhan reported between 1 December 2019 and 28 February 2020. Fig. 2: Estimates of age-specific sCFR and susceptibility to symptomatic infection for COVID-19 in Wuhan
a, Estimates of age-specific sCFRs assuming Psym is 0.50 (red), 0.75 (green) and 0.95 (blue). b, Estimates of relative susceptibility to symptomatic infection by age assuming Psym is 0.50 (red), 0.75 (green) and 0.95 (blue). The markers in both panels show the posterior means and the bars show 95% credible intervals (CrIs). Fig. 3: Estimates of key epidemiologic parameters of the COVID-19 epidemic in Wuhan.
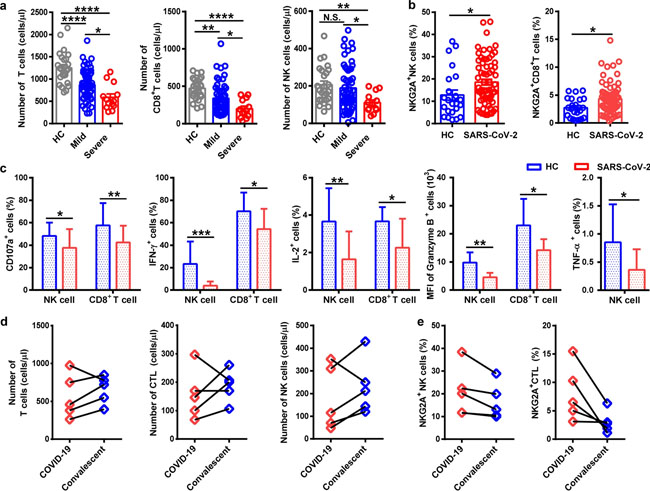
Estimates of basic reproductive number, mean serial interval, initial doubling time, intervention effectiveness, ascertainment rate and the mean time from onset to death, assuming Psym is 0.50 (red), 0.75 (green) and 0.95 (blue). The markers show the posterior means and the bars show 95% CrIs. Correspondence | 19 March 2020 | OPEN 중국 연구 논문: 코로나19 환자에서 항바이러스 림프구의 기능 소진 Functional exhaustion of antiviral lymphocytes in COVID-19 patients 코로나19 감염 환자에서 NK세포가 정상인에 비해 현격히 감소했고, NK세포의 활성을 억제하는 수용체 (NKG2A)의 발현이 높게 나타나는 것을 확인함. 따라서 감염 초기에 면역 세포들의 결손을 방지하고 NK세포의 면역 반응을 개선하는 것이 중요함. 환자들의 NK세포 활성도를 올려줄 경우 유의미한 치료 효과를 볼 수 있을 것으로 예상함. https://www.nature.com/articles/s41423-020-0402-2 Fig. 1
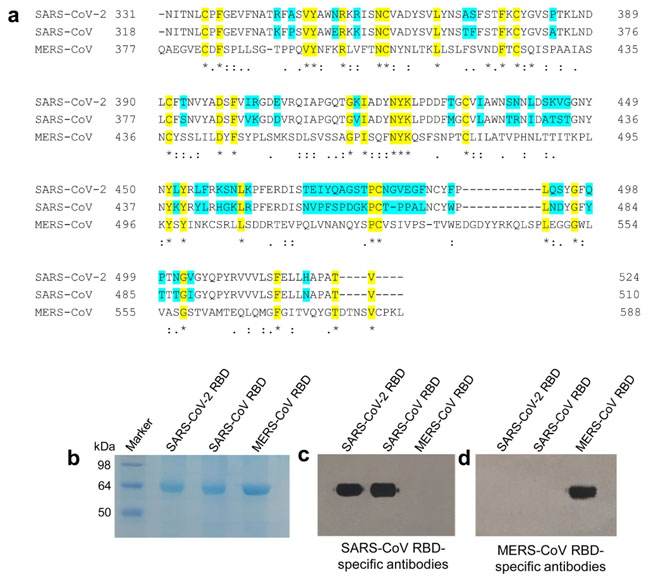
NKG2A+ cytotoxic lymphocytes are functionally exhausted in COVID-19 patients. a Absolute number of T cells, CD8+ T cells, and NK cells in the peripheral blood of healthy controls (n = 25) and patients with mild (n = 55) and severe (n = 13) infection with SARS-CoV-2. b Percentages of NKG2A+ NK cells and NKG2A+CD8+ T cells in the peripheral blood of healthy controls (n = 25) and patients infected with SARS-CoV-2 (n = 68). c Expression of intracellular CD107a, IFN-γ, IL-2, and granzyme-B in gated NK cells and CD8+ T cells and percentage of TNF-α+ NK cells in the peripheral blood of patients infected with SARS-CoV-2 and healthy controls. d Total number of T cells, CTLs, and NK cells in the peripheral blood of COVID-19 patients and convalescing patients. e Percentages of NKG2A+ NK cells and NKG2A+ CTL in the peripheral blood of COVID-19 patients and convalescing patients. Data are mean ± SEM. Unpaired/paired two-tailed Student’s t tests were conducted. p < 0.05 was considered significant. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; N.S., not significant Research | 19 March 2020 2019 신종 코로나바이러스의 수용체 결합 부위 (Receptor Binding Domain, RBD)의 특징 : 바이러스 부착 억제제 및 백신으로서의 RBD 단백질 개발 시사점 Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: implication for development of RBD protein as a viral attachment inhibitor and vaccine https://www.nature.com/articles/s41423-020-0400-4 Fig. 1
Characterization of SARS-CoV-2 RBD. a Multiple sequence alignment of RBDs of SARS-CoV-2, SARS-CoV, and MERS-CoV spike (S) proteins. GenBank accession numbers are QHR63250.1 (SARS-CoV-2 S), AY278488.2 (SARS-CoV S), and AFS88936.1 (MERS-CoV S). Variable amino acid residues between SARS-CoV-2 and SARS-CoV are highlighted in cyan, and conserved residues among SARS-CoV-2, SARS-CoV, and MERS-CoV are highlighted in yellow. Asterisks represent fully conserved residues, colons represent highly conserved residues, and periods represent lowly conserved residues. The alignment was performed using Clustal Omega. SDS-PAGE (b) and Western blot (c, d) analysis of SARS-CoV-2 RBD. The protein molecular weight marker (kDa) is indicated on the left. SARS-CoV and MERS-CoV RBDs were included as controls. Antisera (1:3,000 dilution) from mice immunized with SARS-CoV RBD (c) and MERS-CoV RBD (d) were used for Western blot analysis Fig. 2
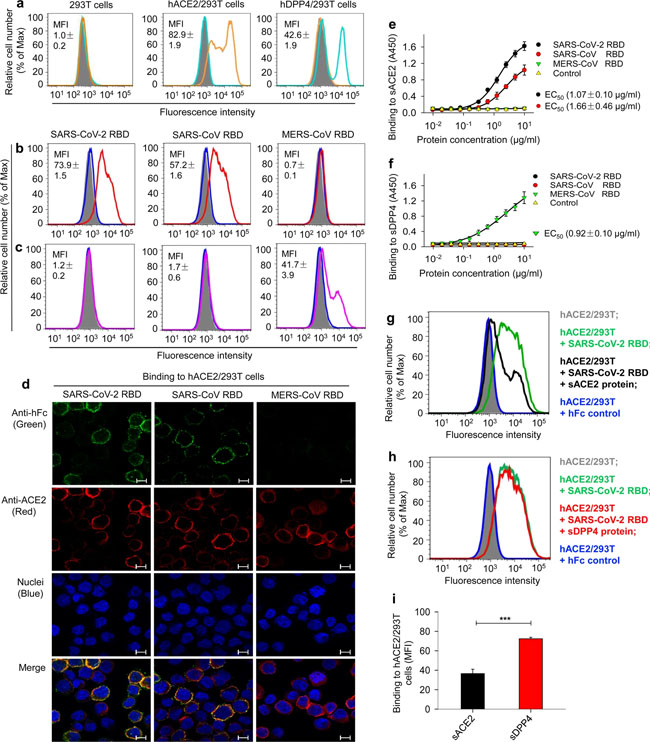
Detection of SARS-CoV-2 RBD binding to human ACE2 receptor. a Flow cytometry analysis of receptor expression in stable cell lines. (left panel) 293T cells alone expressed neither human ACE2 (hACE2) receptor (orange line) nor hDPP4 receptor (cyan line); (middle panel) hACE2-expressing 293T (hACE2/293T) cells expressed only hACE2 (orange line), but not hDPP4 (cyan line); (right panel) hDPP4-expressing 293T (hDPP4/293T) cells expressed only hDPP4 (cyan line), but not hACE2 (orange line). Mock-incubated cells (gray shading) were used as control. Representative images and median fluorescence intensity (MFI) ± standard error (s.e.m.) were shown (n = 4). b, c Flow cytometry analysis of SARS-CoV-2 RBD binding to cell-associated hACE2 receptor in hACE2/293T stable cell lines. SARS-CoV-2 RBD protein bound strongly to hACE2/293T cells (b (left panel, red line)), but not to hDPP4/293T cells (c (left panel, violet line)). SARS-CoV RBD protein bound to hACE2/293T cells (b (middle panel, red line)), but not to hDPP4/293T cells (c (middle panel, violet line)). MERS-CoV RBD protein did not bind to hACE2/293T cells (b (right panel, red line)), but rather bound to hDPP4/293T cells (c (right panel, violet line)). Human IgG Fc (hIgG-Fc, hereinafter hFc) protein-incubated cells (blue line) and mock-incubated cells (gray shading) were included as controls (b, c). Representative images and MFI ± s.e.m. were shown (n = 4). d Immunofluorescence detection of SARS-CoV-2 RBD binding to cell-associated hACE2 receptor in hACE2/293T cells. SARS-CoV-2 RBD protein (green) and SARS-CoV RBD protein (green), each of which was fused with a C-terminal hFc, were stained with FITC-labeled goat anti-human IgG antibody (1:500). hACE2 was stained with a goat-anti-hACE2 antibody (5 μg/ml) and Alexa-Fluor 647-labeled anti-goat antibody (red) (1:200). Fc-fused MERS-CoV RBD protein did not bind to hACE2, so only hACE2 (red), but not RBD (green), was detected in hACE2/293T cells. Nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI, blue). Scale bar: 10 μm. Representative images are shown. e Detection of dose-dependent binding of SARS-CoV-2 RBD protein to soluble hACE2 (sACE2) receptor by ELISA. The SARS-CoV-2 RBD binding to soluble hDPP4 (sDPP4) receptor (f), and the binding of both SARS-CoV RBD and MERS-CoV RBD proteins to sACE2 (e), or sDPP4 (f), were tested. Control: hFc protein. Data are presented as mean A450 ± s.e.m. (n = 4). 50% effective dose (EC50) was calculated for the binding between SARS-CoV-2 RBD (black) or SARS-CoV RBD (red) and hACE2 protein (e, sACE2), or the binding between MERS-CoV RBD and hDPP4 protein (sDPP4, green) (f). g–i Flow cytometry analysis of inhibition of SARS-CoV-2 RBD protein binding to hACE2/293T cells by sACE2. Binding of SARS-CoV-2 RBD to hACE2/293T cells (g, h, green line) was blocked by sACE2 (g, black line), but not by sDPP4 (h, red line). hFc protein-incubated cells (blue line) and mock-incubated cells (gray shading) were included as controls (g, h). Representative images are shown. i The blocking ability of sACE2 or sDPP4, as described above, was expressed as MFI ± s.e.m. (n = 4). Low MFI correlates with high blockage. Experiments were repeated twice and yielded similar results Fig. 3
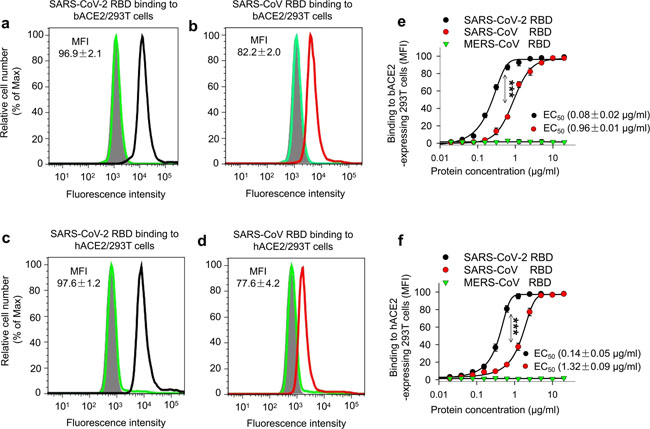
Comparison of SARS-CoV-2 RBD protein binding to human and bat ACE2 receptors. Flow cytometry analysis of SARS-CoV-2 RBD binding to hACE2 and bat ACE2 (bACE2) receptors in 293T cells transiently expressing hACE2 or bACE2. 293T cells were transiently transfected with hACE2 or bACE2 plasmid and incubated with SARS-CoV-2 RBD protein at various concentrations for analysis. SARS-CoV RBD and MERS-CoV RBD proteins were used as controls. Representative images of SARS-CoV-2 RBD protein (2.5 μg/ml) binding to bACE2/293T (a, black line), or hACE2/293T (c, black line), cells were shown. Binding of SARS-CoV RBD protein (2.5 μg/ml) to bACE2/293T (b, red line), or hACE2/293T (d, red line), cells were used as a comparison. MERS-CoV RBD protein (green line) and mock-incubated (gray shading) cells (a–d) were included as controls. e, f Dose-dependent binding of SARS-CoV-2 RBD protein to bACE2/293T (e), or hACE2/293T (f), cells by flow cytometry analysis. Significant differences between binding of SARS-CoV-2 RBD (black) and SARS-CoV RBD (red) to cell-associated bACE2 receptor (e), or hACE2 receptor (f) were identified based on the EC50 values. The data are presented as mean ± s.e.m. (n = 4). Experiments were repeated twice and yielded similar results Fig. 4
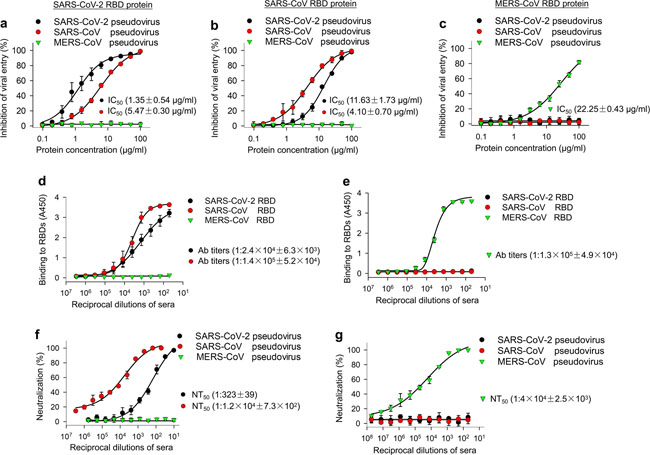
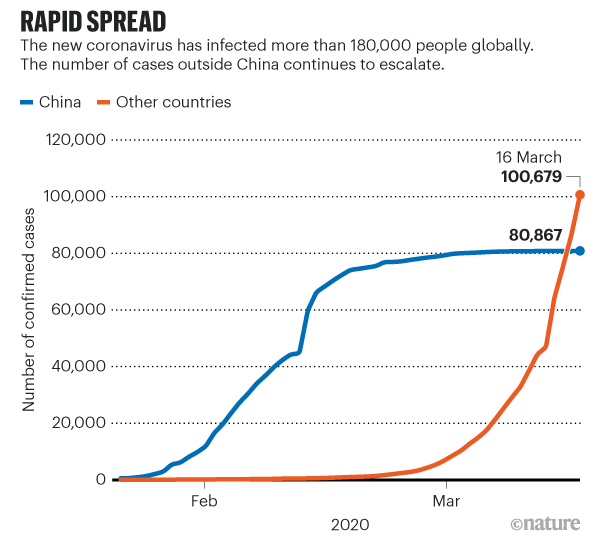
Ability of SARS-CoV-2 RBD to inhibit viral entry, as well as its cross-reactivity and cross-neutralizing activity with SARS-CoV. a Dose-dependent inhibition of SARS-CoV-2 RBD protein against pseudotyped SARS-CoV-2 entry into hACE2/293T cells. SARS-CoV and MERS-CoV RBDs, as well as hDPP4/293T cells, were included as controls. SARS-CoV-2 RBD protein inhibited entry of SARS-CoV-2 and SARS-CoV pseudoviruses into their respective target (hACE2/293T) cells (a), but not the entry of MERS-CoV pseudovirus into its target (hDPP4/293T) cells (a). SARS-CoV RBD protein inhibited both SARS-CoV-2 and SARS-CoV pseudovirus entry, but not MERS-CoV pseudovirus entry (b). MERS-CoV RBD inhibited neither SARS-CoV-2 nor SARS-CoV pseudovirus entry, but it did inhibit MERS-CoV pseudovirus entry (c). The data are presented as mean inhibition (%) ± s.e.m. (n = 4), and 50% inhibition concentration (IC50) was calculated for SARS-CoV-2 RBD (a, b, black), or SARS-CoV RBD (a, b, red), protein against SARS-CoV-2 pseudovirus and SARS-CoV pseudovirus and for MERS-CoV RBD protein (green) against MERS-CoV pseudovirus (c). d Cross-reactivity of SARS-CoV-2 RBD protein with SARS-CoV RBD-specific mouse sera by ELISA. Sera of mice immunized with mammalian cell-expressed SARS-CoV RBD protein30 were tested. Sera of mice immunized with mammalian cell-expressed MERS-CoV RBD protein31 were used as control. The data are presented as mean A450 ± s.e.m. (n = 4). The IgG antibody (Ab) titers were calculated as the endpoint dilution that remains positively detectable for SARS-CoV-2 RBD (black), or SARS-CoV RBD (red), binding to anti-SARS-CoV RBD sera (d) and for MERS-CoV RBD (green) binding to anti-MERS-CoV RBD sera (e). f Cross-neutralization of SARS-CoV RBD-immunized mouse sera against SARS-CoV-2 infection by pseudovirus neutralization assay. MERS-CoV RBD-immunized mouse sera were used as control. The data are presented as mean neutralization (%) ± s.e.m. (n = 4). 50% neutralizing antibody titers (NT50) were calculated against SARS-CoV-2 pseudovirus (black), or SARS-CoV pseudovirus (red), (f) infection in hACE2/293T target cells, as well as against MERS-CoV pseudovirus (green) (g) infection in hDPP4/293T cells. Experiments were repeated twice and yielded similar results News | 18 March 2020 코로나바이러스 팬더믹을 나타내는 5개의 차트 The coronavirus pandemic in five powerful charts https://www.nature.com/articles/d41586-020-00758-2 How is the coronavirus spreading around the world?
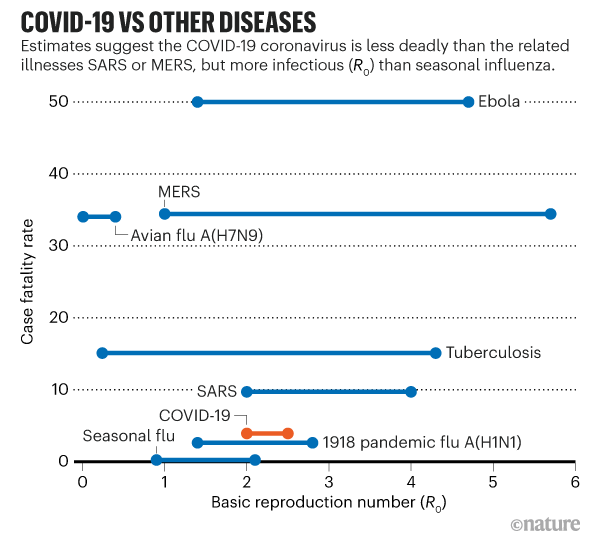
How does COVID-19 compare to other diseases?
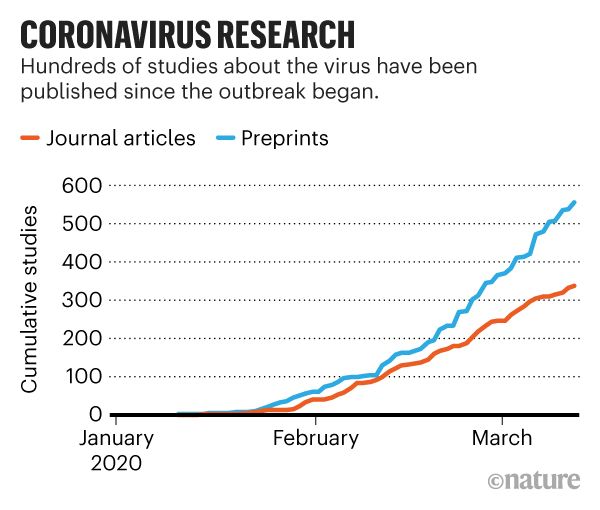
How fast are researchers publishing new coronavirus research?
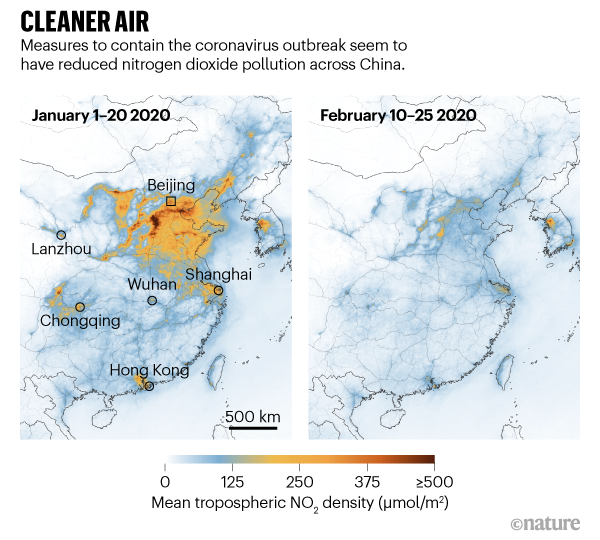
How have travel restrictions affected carbon emissions and air quality?
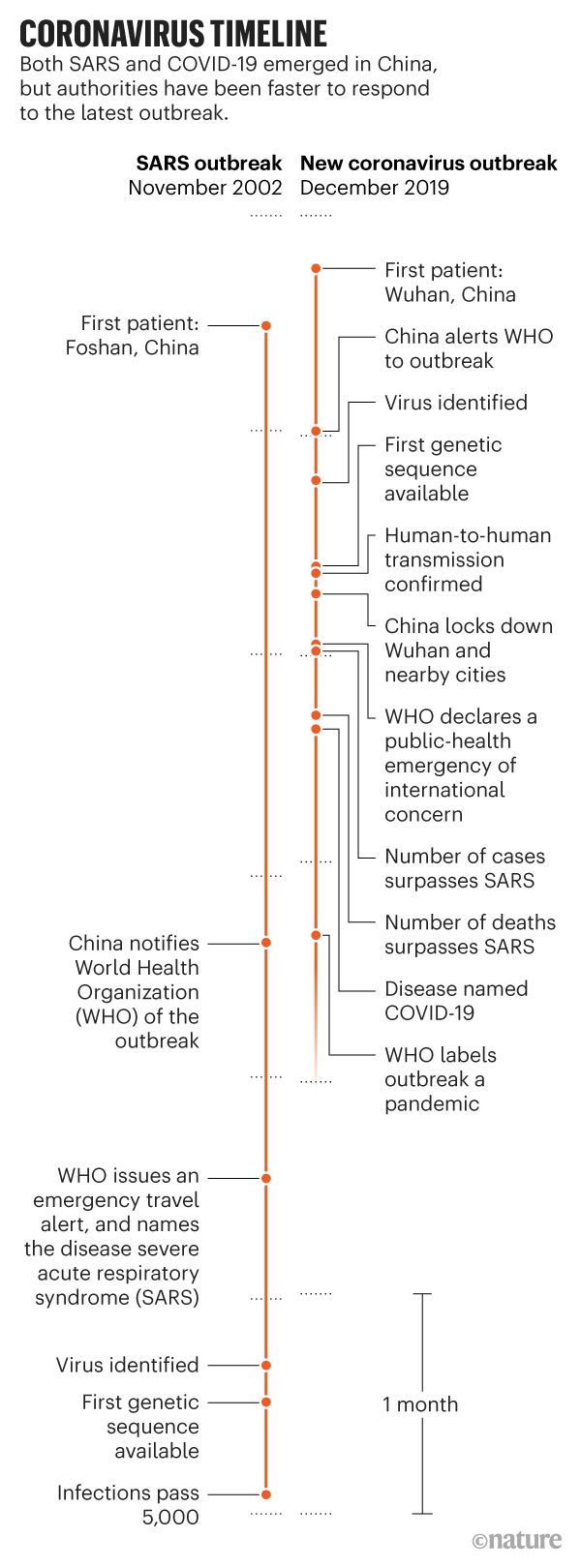
How does the current pandemic compare to the 2003 SARS outbreak?
Correspondence | 18 March 2020 클로로퀸의 독성이 적은 파생 물질인 하이드록시클로로퀸은 생체 외 실험에서 SARS-CoV-2 억제에 효과적이다. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro https://www.nature.com/articles/s41421-020-0156-0 News | 18 March 2020 코로나바이러스 백신: 임상 시험 시작에 있어서 다섯 가지 주요 질문 일부 전문가들은 테스트를 서두르면 위험한 희생이 수반될 것이라고 경고한다. Coronavirus vaccines: five key questions as trials begin Some experts warn that accelerated testing will involve some risky trade-offs. 1) Do people develop immunity? 2) If humans do develop immunity, how long does it last? 3) What kind of immune response should vaccine developers look for? 4) How do we know if a vaccine is likely to work? 5) Will it be safe? https://www.nature.com/articles/d41586-020-00798-8 |